
Calcium signalling in mitochondria and insulin/leptin signalling during ageing
Research summary:
Our interests are understanding calcium regulation of mitochondrial function by way of the calcium-dependent mitochondrial carriers of aspartate-glutamate/AGCs, components of the malate aspartate shuttle (MAS), or ATP.Mg2+-Pi/SCaMCs. These carriers have Ca2+-binding motifs facing the intermembrane space and are not activated by matrix calcium. We also aim at learning the role of these carriers in health and disease.
In neurons, calcium is thought to regulate neuronal activation, by adjusting ATP production to ATP consumption. This occurs thanks to stimulation of glycolysis and OXPHOS. The mitochondrial calcium uniporter (MCU) was thought to play a major role by increasing mitochondrial calcium and OXPHOS in response to activation. We have tested this possibility in neurons using glucose and have found that MCU is dispensable for the increase in respiration in response to neuronal stimulation. Instead, using intracellular sensors of glucose, pyruvate and lactate, we find that Aralar-MAS is required to stimulate glycolysis, pyruvate production and respiration, revealing a calcium dependent mechanism essential to boost glycolysis and respiration in neurons using glucose. Our aim is to study the role of citrin/AGC2 in liver in the mitochondrial response to Ca2+ mobilizing agonists.
Deficiency in Aralar/AGC1 is a rare disease with impaired neurodevelopment, epilepsy and hypomyelination. We have explored treatments for Aralar deficiency and found that β-hydroxybutyrate (βOHB), the main metabolic product of ketogenic diets, is able to overcome the defect in basal and workload-stimulated respiration in Aralar-deficient neurons and partially reverts their failure to produce aspartate and NAA. In vivo administration of βOHB to Aralar-KO mice increases myelin protein levels and dopaminergic markers in these mice, suggesting βHB administration as a potential treatment in Aralar deficiency.
However, whether the defect in myelination of the Aralar-KO mouse is due to the lack of aralar in neurons or oligodendrocytes is unclear. Our present aim is to generate neuron or oligodendrocyte-specific Aralar KO to address these issues.
Citrin deficiency is a urea cycle disorder with different manifestations. Citrin/AGC2 is mainly expressed in liver. In the frame of the Citrin Foundation, we are exploring the exogenous expression of Aralar, which has low expression in normal liver, as possible therapy for Citrin deficiency. We have generated Citrin-KO mice carrying liver-specific Aralar transgene and are studying the effect of the transgene in recovering liver MAS activity and other traits of Citrin deficiency reproduced in Citrin-KO mice.
An ongoing COVID-19 project (CvK), is aimed at developing a therapy against COVID-19 symptoms with the use of senolytics, which would selectively eliminate organisms’ senescent cells.
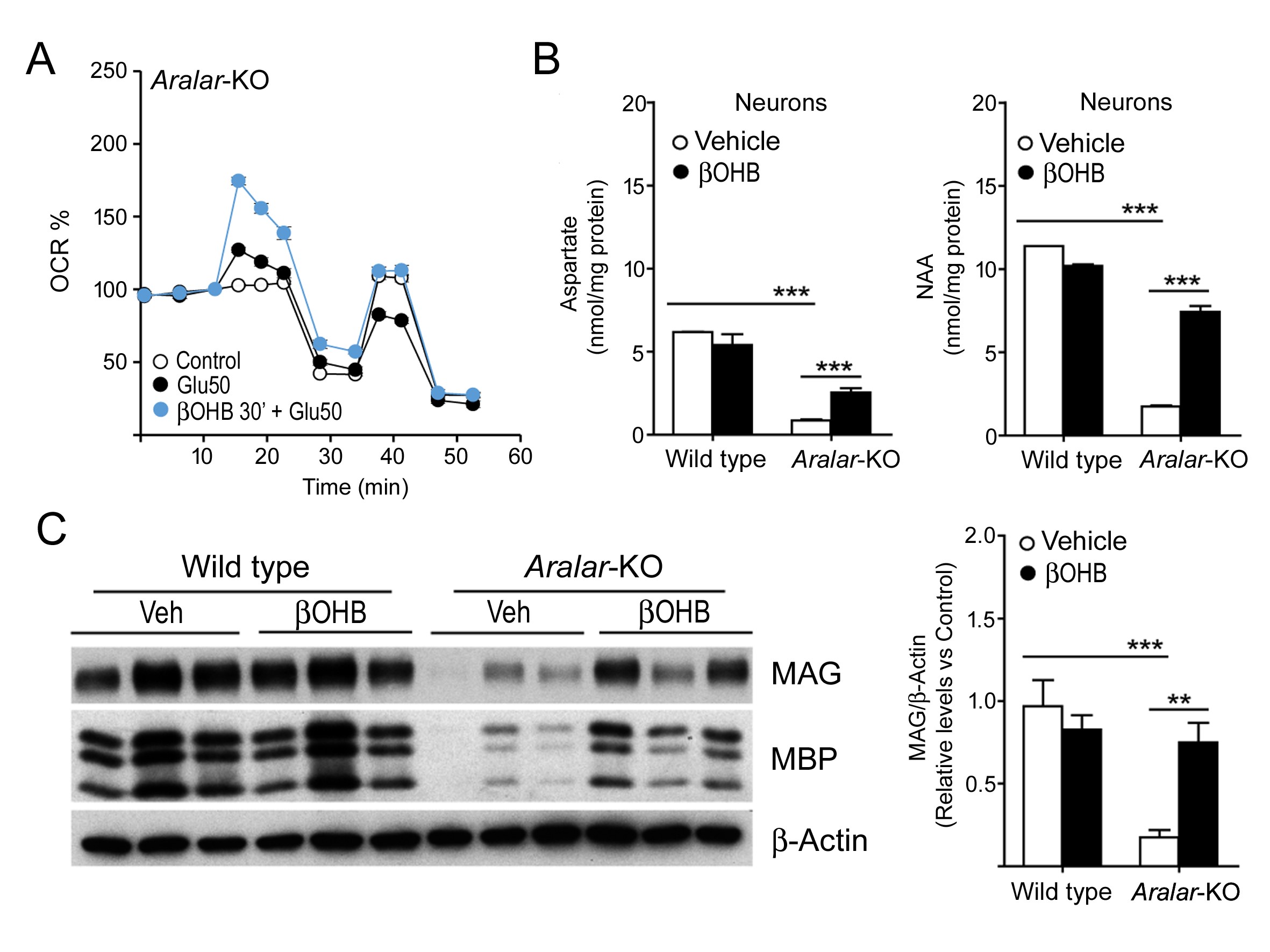
Figure. βOHB Protective roles in Aralar-KO Neurons and Brain: An Alternative to Ketogenic Diet. βOHB increases glutamate-stimulated respiration (A) and the content of Aspartate and NAA (B) in Aralar-KO neuronal cultures, and the levels of cortical myelin proteins in Aralar-KO mice (C) when administered in vivo.

| Lastname | Name | Laboratory | Ext.* | Professional category | |
|---|---|---|---|---|---|
| Bogónez Peláez | Elena | 321 | 4622 | ebogonez(at)cbm.csic.es | Profesor Titular Universidad, GA |
| Carrascosa Baeza | José Mª | 321 | 4651 | jmcarrascosa(at)cbm.csic.es | Catedrático Universidad, GA |
| Contreras Balsa | Laura | 321 | 4651 | lcontreras(at)cbm.csic.es | Titulado Sup.de Actividades Técn. y Profes. GP1 |
| Del Arco | Araceli | 321 | 4651 | adelarco(at)cbm.csic.es | Prof Titular Universidad (convenio UAM-UCLM) |
| González Moreno | Luis | 321 | 4651 | lgmoreno(at)cbm.csic.es | Contrato Predoctoral |
| Juaristi Santos | Inés | 321 | 4651 | ijuaristi(at)cbm.csic.es | Investigador |
| Pardo Merino | Beatriz | 321 | 4635 | bpardo(at)cbm.csic.es | Profesor Titular Universidad, GA |
| Richter | Emma | 321 | 4651 | Becario Erasmus | |
| Romeral Buzón | Alejandro | 4651 | Estudiante TFG | ||
| Santamaría Cano | Andrea | 321 | 4651 | asantamaria(at)cbm.csic.es | Contrato Predoctoral |
| Satrústegui Gil-Delgado | Jorgina | 321 | 4621 | jsatrustegui(at)cbm.csic.es | Emérito |
| Sesé Cobos | Bárbara | 321 | 4651 | bsese(at)cbm.csic.es | E.Ayudantes De Invest. De Los Oo.Publicos De Investigacion |
| Von-Kobbe Alonso | Cayetano-Alfonso | 114.4 | 4809 | cvonkobbe(at)cbm.csic.es |
E.Científicos Titulares de Organismos Públicos de Investigación |
* For external calls please dial 34 91196 followed by the extension number
Relevant publications:
- Pérez-Liébana I, Juaristi I, González-Sánchez P, González-Moreno L, Rial E, Podunavac M, Zakarian A, Molgó J, Vallejo-Illarramendi A, Mosqueira-Martín L, Lopez de Munain A, Pardo B, Satrústegui J, Del Arco A. A Ca2+-Dependent Mechanism Boosting Glycolysis and OXPHOS by Activating Aralar-Malate-Aspartate Shuttle, upon Neuronal Stimulation. J Neurosci. 2022 May 11;42(19):3879-3895. doi: 10.1523/JNEUROSCI.1463-21.2022. Epub 2022 Apr 6. PMID: 35387872; PMCID: PMC9097769.
- Sánchez-González C, Herrero Martín JC, Salegi Ansa B, Núñez de Arenas C, Stančič B, Pereira MP, Contreras L, Cuezva JM, Formentini L. Chronic inhibition of the mitochondrial ATP synthase in skeletal muscle triggers sarcoplasmic reticulum distress and tubular aggregates. Cell Death Dis. 2022 Jun 22;13(6):561. doi: 10.1038/s41419-022-05016-z. PMID: 35732639; PMCID: PMC9217934.
- Pardo B, Herrada-Soler E, Satrústegui J, Contreras L, Del Arco A. AGC1 Deficiency: Pathology and Molecular and Cellular Mechanisms of the Disease. Int J Mol Sci. 2022 Jan 4;23(1):528. doi: 10.3390/ijms23010528. PMID: 35008954; PMCID: PMC8745132.
- Nuevo-Tapioles C, Santacatterina F, Sánchez-Garrido B, de Arenas CN, Robledo-Bérgamo A, Martínez-Valero P, Cantarero L, Pardo B, Hoenicka J, Murphy MP, Satrústegui J, Palau F, Cuezva JM. Effective therapeutic strategies in a preclinical mouse model of Charcot-Marie-Tooth disease. Hum Mol Genet. 2021 Nov 30;30(24):2441-2455. doi: 10.1093/hmg/ddab207. PMID: 34274972; PMCID: PMC8643506.
- Lunetti P, Marsano RM, Curcio R, Dolce V, Fiermonte G, Cappello AR, Marra F, Moschetti R, Li Y, Aiello D, Del Arco Martínez A, Lauria G, De Leonardis F, Ferramosca A, Zara V, Capobianco L. (2021) The mitochondrial aspartate/glutamate carrier (AGC or Aralar1) isoforms in D. melanogaster: biochemical characterization, gene structure, and evolutionary analysis. Biochim Biophys Acta Gen Subj. 1865:129854.
- Esparza-Moltó PB, Romero-Carramiñana I, Núñez de Arenas C, Pereira MP, Blanco N, Pardo B, Bates GR, Sánchez-Castillo C, Artuch R, Murphy MP, Esteban JA, Cuezva JM. (2021) Generation of mitochondrial reactive oxygen species is controlled by ATPase inhibitory factor 1 and regulates cognition. PLoS Biol. 19:e3001252.
- García-Catalán S, González-Moreno L, Del Arco A. (2021) Ca2+-regulated mitochondrial carriers of ATP-Mg2+/Pi: Evolutionary insights in protozoans. Biochim Biophys Acta Mol Cell Res. 1868:119038.
- Rubio C, Lizárraga E, Álvarez-Cilleros D, Pérez-Pardo P, Sanmartín-Salinas P, Toledo-Lobo MV, Alvarez C, Escrivá F, Fernández-Lobato M, Guijarro LG, Valverde AM, Carrascosa JM. (2020) Aging in male Wistar rats associates with changes in intestinal microbiota, gut structure and cholecystokinin-mediated gut-brain axis function. J Gerontol A Biol Sci Med Sci. 2020 Dec 14:glaa313. doi: 10.1093/gerona/glaa313.
- Civera-Tregón A, Domínguez L, Martínez-Valero P, Serrano C, Vallmitjana A, Benítez R, Hoenicka J, Satrústegui J, Palau F. (2021) Mitochondria and calcium defects correlate with axonal dysfunction in GDAP1-related Charcot-Marie-Tooth mouse model. Neurobiol Dis. 152:105300. doi: 10.1016/j.nbd.2021.
- Pérez-Liébana I, Casarejos MJ, Alcaide A, Herrada-Soler E, Llorente-Folch I, Contreras L, Satrústegui J, Pardo B. (2020) βOHB Protective Pathways in Aralar-KO Neurons and Brain: An Alternative to Ketogenic Diet. J Neurosci. 40:9293-9305. doi: 10.1523/JNEUROSCI.0711-20.2020
- Rubio C, Puerto M, García-Rodríquez JJ, Lu VB, García-Martínez I, Alén R, Sanmartín-Salinas P, Toledo-Lobo MV, Saiz J, Ruperez J, Barbas C, Menchén L, Gribble FM, Reimann F, Guijarro LG, Carrascosa JM, Valverde ÁM. (2020) Impact of global PTP1B deficiency on the gut barrier permeability during NASH in mice. Mol Metab. 35:100954. doi: 10.1016/j.molmet.2020.01.018
- Juaristi I, Llorente-Folch I, Satrústegui J, Del Arco A. (2019) Extracellular ATP and glutamate drive pyruvate production and energy demand to regulate mitochondrial respiration in astrocytes. Glia 67:759-774. doi: 10.1002/glia.23574.
- Puertas-Frías G, Del Arco A, Pardo B, Satrústegui J, Contreras L (2019) Mitochondrial movement in Aralar/Slc25a12/AGC1 deficient cortical neurons. Int. 131:104541. doi: 10.1016/j.neuint.2019.104541.
- von Kobbe C (2019) Targeting senescent cells: approaches, opportunities, challenges. Aging (Albany NY) 11: 12844–12861. doi: 10.18632/aging.102557.
- Remacha L, Comino-Méndez I, Richter S, Contreras L, Currás-Freixes M, Pita G, Letón R, Galarreta A, Torres-Pérez R, Honrado E, Jiménez S, Maestre L, Moran S, Esteller M, Satrústegui J, Eisenhofer G, Robledo M, Cascón A. (2017) Targeted Exome Sequencing of Krebs Cycle Genes Reveals Candidate Cancer-Predisposing Mutations in Pheochromocytomas and Paragangliomas. Clin Cancer Res. 23:6315-6324. doi: 10.1158/1078-0432.CCR-16-2250.
- González-Sánchez P, Del Arco A, Esteban JA, Satrústegui J. (2017) Store-Operated Calcium Entry Is Required for mGluR-Dependent Long Term Depression in Cortical Neurons. Front Cell Neurosci. 11:363. doi: 10.3389/fncel.2017.00363.
- Writzl K, Maver A, Kovačič L, Martinez-Valero P, Contreras L, Satrustegui J, Castori M, Faivre L, Lapunzina P, van Kuilenburg ABP, Radović S, Thauvin-Robinet C, Peterlin B, Del Arco A, Hennekam RC. (2017) De Novo Mutations in SLC25A24 Cause a Disorder Characterized by Early Aging, Bone Dysplasia, Characteristic Face, and Early Demise. Am J Hum Genet. 101:844-855. doi: 10.1016/j.ajhg.2017.09.017.
- Juaristi I, García-Martín ML, Rodrigues TB, Satrústegui J, Llorente-Folch I, Pardo B. (2017) ARALAR/AGC1 deficiency, a neurodevelopmental disorder with severe impairment of neuronal mitochondrial respiration, does not produce a primary increase in brain lactate. J Neurochem. 142:132-139. doi: 10.1111/jnc.14047.
- González-Sánchez P, Pla-Martín D, Martínez-Valero P, Rueda CB, Calpena E, Del Arco A, Palau F, Satrústegui J. (2017) CMT-linked loss-of-function mutations in GDAP1 impair store-operated Ca2+ entry-stimulated respiration. Sci Rep. 7:42993. doi: 10.1038/srep42993.
- Rueda, C.B., Traba, J., Amigo, I., Llorente-Folch, I., González-Sánchez, P., Pardo, B., Esteban, J.A., del Arco, A. and Satrústegui, J. (2015) Mitochondrial ATP-Mg/Pi carrier SCaMC-3/Slc25a23 counteracts PARP-1- dependent fall in mitochondrial ATP caused by excitotoxic insults in neurons. J. Neurosci. 35, 3566-3581.
- Llorente-Folch, I., Rueda, C.B., Pardo, B., Szabadkai, G., Duchen, M.R. and Satrustegui, J. (2015) The regulation of neuronal mitochondrial metabolism by calcium. J. Physiol. 593, 3447-3462.
- Llorente-Folch, I., Rueda, C.B., Perez-Liebana, I., Satrústegui, J. and Pardo, B. (2016) L-Lactate-Mediated Neuroprotection against Glutamate-Induced Excitotoxicity Requires Aralar/AGC1. J. Neurosci. 36, 4443-4456.
- Amigo, I., Traba, J., González-Barroso, MM., Rueda, CB., Fernández, M., Rial, E., Sánchez, A, Satrústegui, J. and del Arco, A. (2013) Glucagon regulation of oxidative phosphorylation requires an increase in matrix adenine nucleotide content through Ca2+ activation of the mitochondrial ATP-Mg/Pi carrier SCaMC-3. J. Biol. Chem. 288, 7791-7802.
- Llorente-Folch, I., Rueda, CB., Amigo, I., del Arco, A., Saheki, T., Pardo, B. and Satrústegui, J. (2013) Calcium-regulation of mitochondrial respiration maintains ATP homeostasis and requires ARALAR/AGC1-malate aspartate shuttle in intact cortical neurons. J. Neurosci. 33, 13957-13971.
- Lindsay, KJ., Du, J., Sloat, SR., Contreras, L., Linton, JD., Turner, SJ., Sadilek, M., Satrústegui, J. and Hurley, JB. (2014) Pyruvate kinase and aspartate-glutamate carrier distributions reveal key metabolic links between neurons and glia in retina. Proc. Natl. Acad. Sci. U. S. A. 111, 15579-15584.
- Llorente-Folch, I., Sahún, I., Contreras, L., Casarejos, MJ., Grau JM, Saheki, T., Mena, MA., Satrústegui, J., Dierssen M. and Pardo, B. (2013) AGC1-malate aspartate shuttle activity is critical for dopamine handling in the nigrostriatal pathway. J. Neurochem. 124, 347-362.
Capítulos de libros:
- del Arco, A. and Satrústegui, J. (March 2013) Mitochondrial Carriers. In: eLS. John Wiley & Sons, Ltd: Chichester.
- José María Carrascosa Baeza, “Ciencia, ética y sostenibilidad: el papel de la Academia para alcanzar los Objetivos de Desarrollo Sostenible” Agenda 2030: Claves para la transformación sostenible / Margarita Alfaro (ed. lit.), Silvia Arias Careaga (ed. lit.), Ana Gamba Romero (ed. lit.), ISBN 9788490977071.
Doctoral theses:
- Irene Pérez Liébana (2020) “The mitochondrial aspartate-glutamate carrier Aralar/AGC1 controls neuronal respiration and (beta)-Hydroxybutyrate rescues brain defects caused by AGC1 deficiency”. UAM. Jorgina Satrústegui y Beatriz Pardo.
- Inés Juaristi Santos (2019) “Regulation of mitochondrial respiration in astrocytes: role of Ca2+, ATP demand and pyruvate production”. UAM. Jorgina Satrústegui y Araceli del Arco.
- Carmen Rubio Caballero (2018) “Changes in gut microbiota associated with inflammation during ageing and non-alcoholic steatohepatitis”. UAM. José María Carrascosa Baeza y Ángela Martínez Valverde.
- Paula Martínez Valero (2017) “Enfermedad de Charcot-Marie-Tooth asociada a Gadp1: alteraciones en nocicepción y señalización por calcio”. UAM. Jorgina Satrústegui, Laura Contreras y Carolina Roza.
- Paloma González Sánchez (2017) “Store-operated calcium entry in neural cells. A role in Charcot-Marie-Tooth disease”. UAM. Jorgina Satrústegui y Araceli del Arco.
- Carlos B. Rueda Díez (2014). “Ca2+ modulation of mitochondrial function under physiological and pathological stimulation: Role of the ATP-Mg/Pi carrier, SCaMC-3”. UAM. Jorgina Satrústegui, Beatriz Pardo y Araceli del Arco.
- Irene Llorente Folch (2013). “New roles of aralar, the brain mitochondrial aspartate/glutamate carrier in dopamine handling glutamate excitotoxicity and regulation of mitochondrial respiration”. UAM. Jorgina Satrústegui y Beatriz Pardo.
- Ignacio Amigo de la Huerga (2013). “Characterization of SCaMC-3, the mitochondrial ATP-Mg/Pi carrier present in liver and brain”. UAM. Jorgina Satrústegui y Araceli del Arco


















