
Molecular mechanisms of Oligodendrocyte-Neuron interaction & pathologies associated with myelin
Research summary:
In our laboratory we study the neurological component of demyelinating pathologies and investigate the molecular mechanisms responsible for the processes of myelination in the Central Nervous System (CNS). Adequate myelination is essential for the correct transmission of the nerve impulse. In the CNS, oligodendrocytes are the cells responsible for myelination of neuronal axons, through a complex process that requires complex cellular interactions. In the absence of a correct myelination, diseases such as Multiple Sclerosis or leukodystrophies appear, currently orphans of an effective treatment. The possibility of generating therapies based on the neurological component of these diseases could stimulate the regeneration of new oligodendrocytes or increase the capacity of the remaining set of oligodendrocytes to produce more myelin and reestablish correct myelination. Our group has demonstrated the importance of the GTPases R-Ras1 and R-Ras2, essential proteins in the differentiation and survival of OLs. Moreover, we have described to the relevance of their presence for the maintenance of energetic homeostasis and for the correct functioning of the nerve impulse transmission. Models lacking R-Ras1 and/or R-Ras2 faithfully reproduce the symptomatological characteristics of myelin diseases and could be used as models for the development of new treatments based on the neurological component.
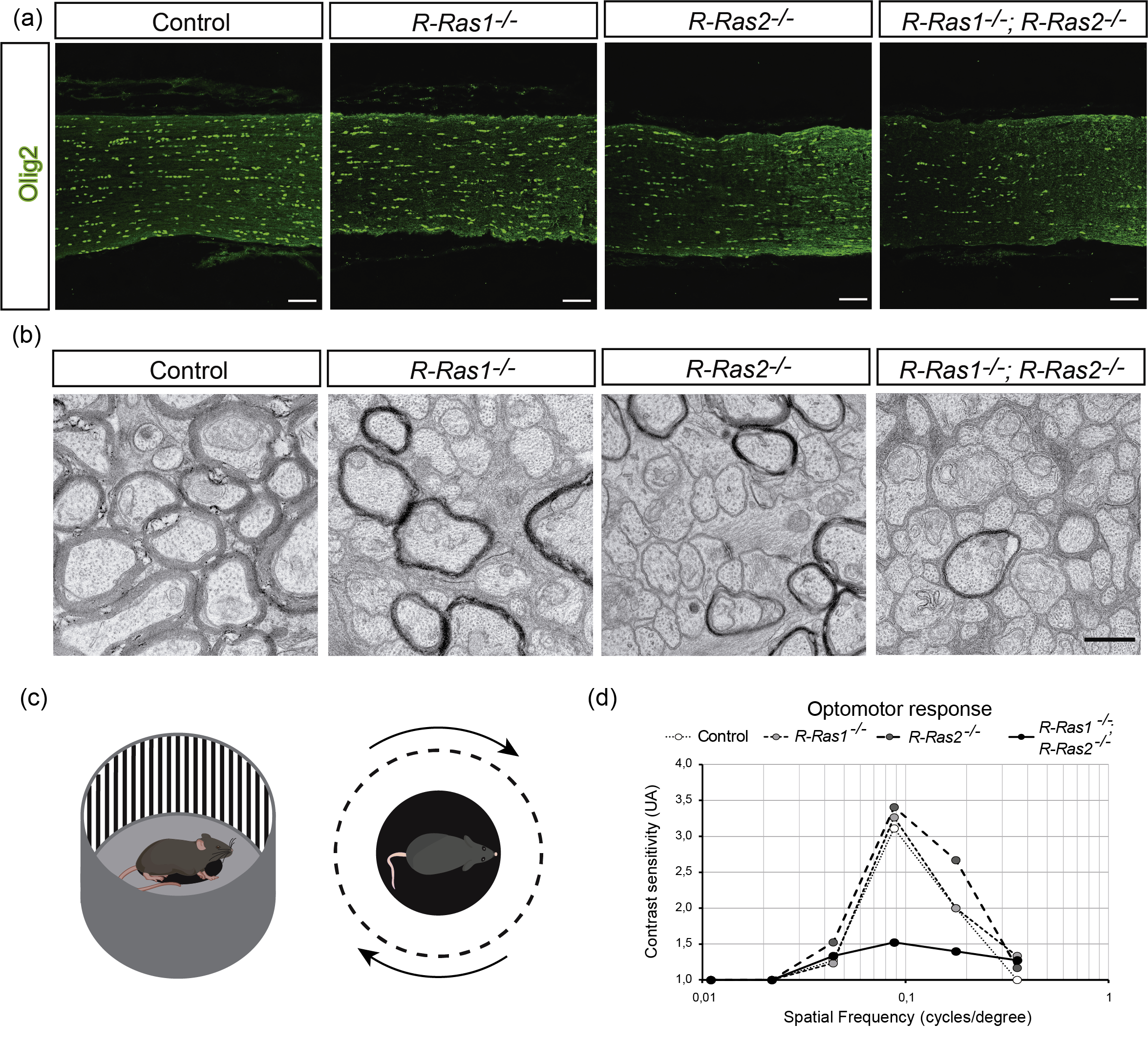
Figure 1.
(a) Loss of oligodendrocytes in mutant mice lacking from R-Ras1, R-Ras2, or both. (b) Axons lose their protective myelin sheath when they lack R-Ras1 and/or R-Ras2. (c) and (d) Loss of visual function generated by the loss of oligodendrocytes and myelin in the mutant mice.

| Last name | Name | Laboratory | Ext.* | Professional category | |
|---|---|---|---|---|---|
| Alcalá Morote | Pablo | 305 | 4650 | Estudiante TFM | |
| Alcover Sánchez | Berta | 305 | 4650 | berta.alcover(at)cbm.csic.es | Titulado Sup.de Actividades Técn. y Profes. GP1 |
| Cubelos Alvarez | Beatriz | 305 | 4650 | bcubelos(at)cbm.csic.es | Profesor Titular Universidad, GA |
Relevant publications:
- Garcia-Martin G., Sanz-Rodriguez M., Alcover-Sanchez B., Pereira MP., Wandosell F., Cubelos B. R-Ras1 and R-Ras2 Expression in Anatomical Regions and Cell Types of the Central Nervous System. Int J Mol Sci. 2022 Jan 17;23(2):978. doi: 10.3390/ijms23020978. PMID: 35055164. (IJMS, IF: 5.92).
- Garcia-Martin G., Alcover-Sanchez B., Wandosell F., Cubelos B. “Pathways involved in remyelination after cerebral ischemia”. Curr Neuropharmacol. 2021 Jun 9. doi: 10.2174/1570159X19666210610093658. Epub ahead of print. PMID: 34151767. (Curr Neuropharmacol., IF: 4.66).
- Alcover-Sanchez B, Garcia-Martin G, Escudero-Ramirez J, Gonzalez-Riano C, Lorenzo P, Gimenez-Cassina A, Formentini L, de la Villa-Polo P, Pereira MP, Wandosell F, Cubelos B. Absence of R-Ras1 and R-Ras2 causes mitochondrial alterations that trigger axonal degeneration in a hypomyelinating disease model. Glia. 2021 Mar;69(3):619-637. doi: 10.1002/glia.23917. Epub 2020 Oct 3. PMID: 33010069.
- Alcover-Sanchez B, Garcia-Martin G, Wandosell F, Cubelos B. R-Ras GTPases Signaling Role in Myelin Neurodegenerative Diseases. Int J Mol Sci. 2020 Aug 17;21(16):5911. doi: 10.3390/ijms21165911. PMID: 32824627; PMCID: PMC7460555.
- Sanz-Rodriguez M, Gruart A, Escudero-Ramirez J, de Castro F, Delgado-García JM, Wandosell F, Cubelos B. R-Ras1 and R-Ras2 Are Essential for Oligodendrocyte Differentiation and Survival for Correct Myelination in the Central Nervous System. J Neurosci. 2018 May 30;38(22):5096-5110. doi: 10.1523/JNEUROSCI.3364-17.2018. Epub 2018 May 2. PMID: 29720552; PMCID: PMC6705947.
- Gonzalez-Riano C, Sanz-Rodríguez M, Escudero-Ramirez J, Lorenzo MP, Barbas C, Cubelos B, Garcia A. Target and untargeted GC-MS based metabolomic study of mouse optic nerve and its potential in the study of neurological visual diseases. J Pharm Biomed Anal. 2018 May 10;153:44-56. doi: 10.1016/j.jpba.2018.02.015. Epub 2018 Feb 10. PMID: 29459235.
- Doan RN, Bae BI, Cubelos B, Chang C, Hossain AA, Al-Saad S, Mukaddes NM, Oner O, Al-Saffar M, Balkhy S, Gascon GG; Homozygosity Mapping Consortium for Autism, Nieto M, Walsh CA. Mutations in Human Accelerated Regions Disrupt Cognition and Social Behavior. Cell. 2016 Oct 6;167(2):341-354.e12. doi: 10.1016/j.cell.2016.08.071. Epub 2016 Sep 22. PMID: 27667684; PMCID: PMC5063026.
- Cubelos B, Briz CG, Esteban-Ortega GM, Nieto M. Cux1 and Cux2 selectively target basal and apical dendritic compartments of layer II-III cortical neurons. Dev Neurobiol. 2015 Feb;75(2):163-72. doi: 10.1002/dneu.22215. Epub 2014 Jul 30. PMID: 25059644.
- Gutierrez-Erlandsson S, Herrero-Vidal P, Fernandez-Alfara M, Hernandez-Garcia S, Gonzalo-Flores S, Mudarra-Rubio A, Fresno M, Cubelos B. R-RAS2 overexpression in tumors of the human central nervous system. Mol Cancer. 2013 Oct 23;12(1):127. doi: 10.1186/1476-4598-12-127. PMID: 24148564; PMCID: PMC3900289.
- Merino JJ, Bellver-Landete V, Oset-Gasque MJ, Cubelos B. CXCR4/CXCR7 molecular involvement in neuronal and neural progenitor migration: focus in CNS repair. J Cell Physiol. 2015 Jan;230(1):27-42. doi: 10.1002/jcp.24695. PMID: 24913264.
- Cubelos B, Leite C, Giménez C, Zafra F. Localization of the glycine transporter GLYT1 in glutamatergic synaptic vesicles. Neurochem Int. 2014 Jul;73:204-10. doi: 10.1016/j.neuint.2013.09.002. Epub 2013 Sep 11. PMID: 24036061.
- Rodríguez-Tornos FM, San Aniceto I, Cubelos B, Nieto M. Enrichment of conserved synaptic activity-responsive element in neuronal genes predicts a coordinated response of MEF2, CREB and SRF. PLoS One. 2013;8(1):e53848. doi: 10.1371/journal.pone.0053848. Epub 2013 Jan 31. PMID: 23382855; PMCID: PMC3561385.
- Cubelos B, Nieto M. Intrinsic programs regulating dendrites and synapses in the upper layer neurons of the cortex. Commun Integr Biol. 2010 Nov;3(6):483-6. doi: 10.4161/cib.3.6.12755. Epub 2010 Nov 1. PMID: 21331220; PMCID: PMC3038044.
- Cubelos B, Sebastián-Serrano A, Beccari L, Calcagnotto ME, Cisneros E, Kim S, Dopazo A, Alvarez-Dolado M, Redondo JM, Bovolenta P, Walsh CA, Nieto M. Cux1 and Cux2 regulate dendritic branching, spine morphology, and synapses of the upper layer neurons of the cortex. 2010 May 27;66(4):523-35. doi: 10.1016/j.neuron.2010.04.038. PMID: 20510857; PMCID: PMC2894581.
- Delgado P, Cubelos B, Calleja E, Martínez-Martín N, Ciprés A, Mérida I, Bellas C, Bustelo XR, Alarcón B. Essential function for the GTPase TC21 in homeostatic antigen receptor signaling. Nat Immunol. 2009 Aug;10(8):880-8. doi: 10.1038/ni.1749. Epub 2009 Jun 28. PMID: 19561613.
- Cubelos B, Sebastián-Serrano A, Kim S, Redondo JM, Walsh C, Nieto M. Cux-1 and Cux-2 control the development of Reelin expressing cortical interneurons. Dev Neurobiol. 2008 Jun;68(7):917-25. doi: 10.1002/dneu.20626. PMID: 18327765; PMCID: PMC2938960.
- Cubelos B, Sebastián-Serrano A, Kim S, Moreno-Ortiz C, Redondo JM, Walsh CA, Nieto M. Cux-2 controls the proliferation of neuronal intermediate precursors of the cortical subventricular zone. Cereb Cortex. 2008 Aug;18(8):1758-70. doi: 10.1093/cercor/bhm199. Epub 2007 Nov 21. PMID: 18033766.
- González-González IM, García-Tardón N, Cubelos B, Giménez C, Zafra F. The glutamate transporter GLT1b interacts with the scaffold protein PSD-95. J Neurochem. 2008 Jun;105(5):1834-48. doi: 10.1111/j.1471-4159.2008.05281.x. Epub 2008 Feb 4. PMID: 18248606.
- Fernández-Sánchez E, Díez-Guerra FJ, Cubelos B, Giménez C, Zafra F. Mechanisms of endoplasmic-reticulum export of glycine transporter-1 (GLYT1). Biochem J. 2008 Feb 1;409(3):669-81. doi: 10.1042/BJ20070533. PMID: 17919119.
- Cubelos B, Giménez C, Zafra F. Localization of the GLYT1 glycine transporter at glutamatergic synapses in the rat brain. Cereb Cortex. 2005 Apr;15(4):448-59. doi: 10.1093/cercor/bhh147. PMID: 15749988.
- Cubelos B, González-González IM, Giménez C, Zafra F. The scaffolding protein PSD-95 interacts with the glycine transporter GLYT1 and impairs its internalization. J Neurochem. 2005 Nov;95(4):1047-58. doi: 10.1111/j.1471-4159.2005.03438.x. PMID: 16271045.
- Cubelos B, Giménez C, Zafra F. The glycine transporter GLYT1 interacts with Sec3, a component of the exocyst complex. 2005 Nov;49(6):935-44. doi: 10.1016/j.neuropharm.2005.07.021. Epub 2005 Sep 21. PMID: 16181645.
- González-González IM, Cubelos B, Giménez C, Zafra F. Immunohistochemical localization of the amino acid transporter SNAT2 in the rat brain. 2005;130(1):61-73. doi: 10.1016/j.neuroscience.2004.09.023. PMID: 15561425.
- Cubelos B, Gonzalez-Gonzalez I, Gimenez C and Zafra F. The amino acid transporter SN2 (SLC38A5) localizes to glial in the rat brain. Glia 2004; 49(2): 230-244
- Nogal ML, González de Buitrago G, Rodríguez C, Cubelos B, Carrascosa AL, Salas ML, Revilla Y. African swine fever virus IAP homologue inhibits caspase activation and promotes cell survival in mammalian cells. J Virol. 2001 Mar;75(6):2535-43. doi: 10.1128/JVI.75.6.2535-2543.2001. PMID: 11222676; PMCID: PMC115875.
- Cubelos B, Gimenéz C y Zafra F. Hipótesis glutamatérgica de la esquizofrenia: Mecanismos moleculares del transporte de glicina en las sinapsis glutamatérgicas.Real Academia Nacional Farmacia Volumen:Nº41; 5-25. ISSN 0034-0618
Tesis doctorales:
- Berta Alcover Sánchez (Estudiante de Doctorado. Universidad Autónoma de Madrid) en curso.
- Gonzalo García Martín (Estudiante de Doctorado. Universidad Autónoma de Madrid) en curso.
- Brian Ernie Quinones (Estudiante de Doctorado. Universidad Autónoma de Madrid) en curso.
- Miriam Sanz Rodríguez. Tesis doctoral: “Papel de R-Ras1 y R-Ras2 en los procesos de mielinización del SNC”. Universidad Autónoma de Madrid. Junio 2018. Calificación: Sobresaliente “Cum Laude”.
Trabajos de fin de grado y máster:
- Rosa Plaza Clavero (Estudiante de Máster. Universidad Autónoma de Madrid) en curso.
- Gonzalo García Martín. Trabajo de Fin de Máster: “Role of R-Ras on the Oligodendrocyte actin cytoskeleton”. Universidad Autónoma de Madrid 2021.
- Jorge Navarro Nadal. Trabajo de Fin de Máster. Universidad Complutense de Madrid 2021.
- Juan Escudero Ramírez. Trabajo de Fin de Máster: “Papel de RRas1 y RRas2 en la función mitocondrial del SNC”. Universidad Autónoma de Madrid. 2019.
- Juan Ramón Perea Úbeda-Portugués. Trabajo de Fin de Máster: “Papel de RRas2 en los procesos de mielinización en el nervio óptico”. Autónoma de Madrid. 2015. Premio Especial de la Fundación ONCE al mejor trabajo de investigación curso 2014/ 2015. Accésit en el Certamen Arquímedes al mejor trabajo original de investigación. BOE Núm. 289; 3 de diciembre de 2015.
- Andrea Alcaraz Ramírez (Estudiante de Grado. Universidad Autónoma de Madrid) en curso.
- Gonzalo García Martín. Trabajo de Fin de Grado: “El déficit de mielina desencadena degeneración axonal en un modelo de ratón hipomielinizante”. Universidad Autónoma de Madrid. 2020.
- Germán Benito Vicente. Trabajo de Fin de Grado: “Estudio de la contribución de R-Ras1 y R-Ras2 en la morfología oligodendrocitaria”. Universidad Autónoma de Madrid. 2019.
- Juan Escudero Ramírez. Trabajo de Fin de Grado: “Papel de RRas1 y RRas2 en el Sistema Nervioso”. Universidad Autónoma de Madrid. 2017.
- Manuel Troncoso. Trabajo de Fin de Grado: “Papel de RRas1 y RRas2 la mielinización del nervio óptico de ratones”. Universidad Autónoma de Madrid. 2017.
- Pedro Herrero Vidal. Trabajo de Fin de Grado: “Papel de RRas2 en el desarrollo y función de la retina”. Universidad Autónoma de Madrid. 2014.
- Juan Ramón Perea Úbeda-Portugués. Trabajo de Fin de Grado: “Papel de RRas2 la función de la retina”. Universidad Autónoma de Madrid. 2013.
- Raquel Forcén García. Trabajo de Fin de Grado: “Papel de RRas2 en el desarrollo de la retina”. Universidad Autónoma de Madrid. 2013.
- Raquel Martín Morales. Trabajo de Fin de Grado: “Papel de RRas2 en el sistema nervioso”. Universidad Autónoma de Madrid. 2012.


















