
Señalización mitocondrial del calcio y señalización de insulina/leptina en envejecimiento
Resumen de Investigación:
El interés de nuestra investigación reside en comprender la regulación por calcio de la actividad mitocondrial mediada por los transportadores mitocondriales de aspartato/glutamato (AGCs), componentes de la lanzadera de malato/aspartato (MAS) o de ATP-Mg2+/Pi (SCaMCs). Estos transportadores presentan dominios de unión a calcio orientados hacia el espacio intermembrana y que no son activados por el calcio de la matriz mitocondrial. También nuestro objetivo es conocer el papel fisiológico de estos transportadores y sus implicaciones patológicas.
En las neuronas, se considera que el calcio regula la activación neuronal ajustando la producción de ATP al consumo de ATP. Esto ocurre gracias a la estimulación de la glucólisis y los procesos de fosforilación oxidativa (OXPHOS). Se creía que el uniportador de calcio mitocondrial (MCU) desempeñaba un papel importante al aumentar el calcio mitocondrial y la actividad OXPHOS en respuesta a la activación. Hemos probado esta posibilidad y hemos descubierto que MCU no se requiere para aumentar la respiración mitocondrial en respuesta a la estimulación en neuronas que utilizan glucosa. En cambio, utilizando sensores intracelulares de glucosa, piruvato y lactato, encontramos que se requiere Aralar/AGC1-MAS para estimular la glucólisis, la producción de piruvato y la respiración, indicando que representan un mecanismo dependiente de calcio esencial para estimular la glucólisis y la respiración en las neuronas usando glucosa. Nuestro objetivo actual es estudiar el papel de Citrin/AGC2 en el hígado en la respuesta mitocondrial a agonistas movilizadores de Ca2+.
La deficiencia de Aralar es una enfermedad rara que cursa con deterioro del neurodesarrollo, epilepsia e hipomielinización. Hemos explorado tratamientos para esta enfermedad y hemos descubierto que el β-hidroxibutirato (βOHB), el principal producto metabólico de las dietas cetogénicas, es capaz de superar el defecto en la respiración, tanto basal como estimulada por cargas de trabajo, en neuronas con deficiencia de Aralar/AGC1 y que revierte parcialmente su incapacidad para producir aspartato y NAA. La administración in vivo de βOHB a ratones Aralar-KO aumenta los niveles de proteínas de mielina y de marcadores dopaminérgicos en estos ratones, sugiriendo la administración de βOHB como un tratamiento potencial para la deficiencia de Aralar/AGC1.
Sin embargo, no está claro si el defecto en la mielinización del ratón Aralar-KO se debe a la falta de Aralar/AGC1 en las neuronas o en los oligodendrocitos. Nuestro objetivo actual es generar ratones Aralar-KO específicos de neuronas u oligodendrocitos para abordar estos problemas.
La deficiencia de Citrin es un trastorno del ciclo de la urea con diferentes manifestaciones clínicas. Citrin/AGC2 se expresa principalmente en el hígado. En el marco de la Fundación Citrin, estamos explorando la expresión exógena de Aralar, que tiene baja expresión en hígado normal, como posible terapia para la deficiencia de Citrin. Hemos generado ratones Citrin-KO que portan un transgén para Aralar/AGC1 con expresión específica de hígado y estamos estudiando su efecto en la recuperación de la actividad MAS del hígado y otros rasgos de la deficiencia de Citrin reproducidos en ratones Citrin-KO.
Un proyecto de COVID-19 (CvK) en curso tiene como objetivo desarrollar una terapia contra los síntomas de COVID-19 con el uso de senolíticos, que eliminarían selectivamente las células senescentes de los organismos.
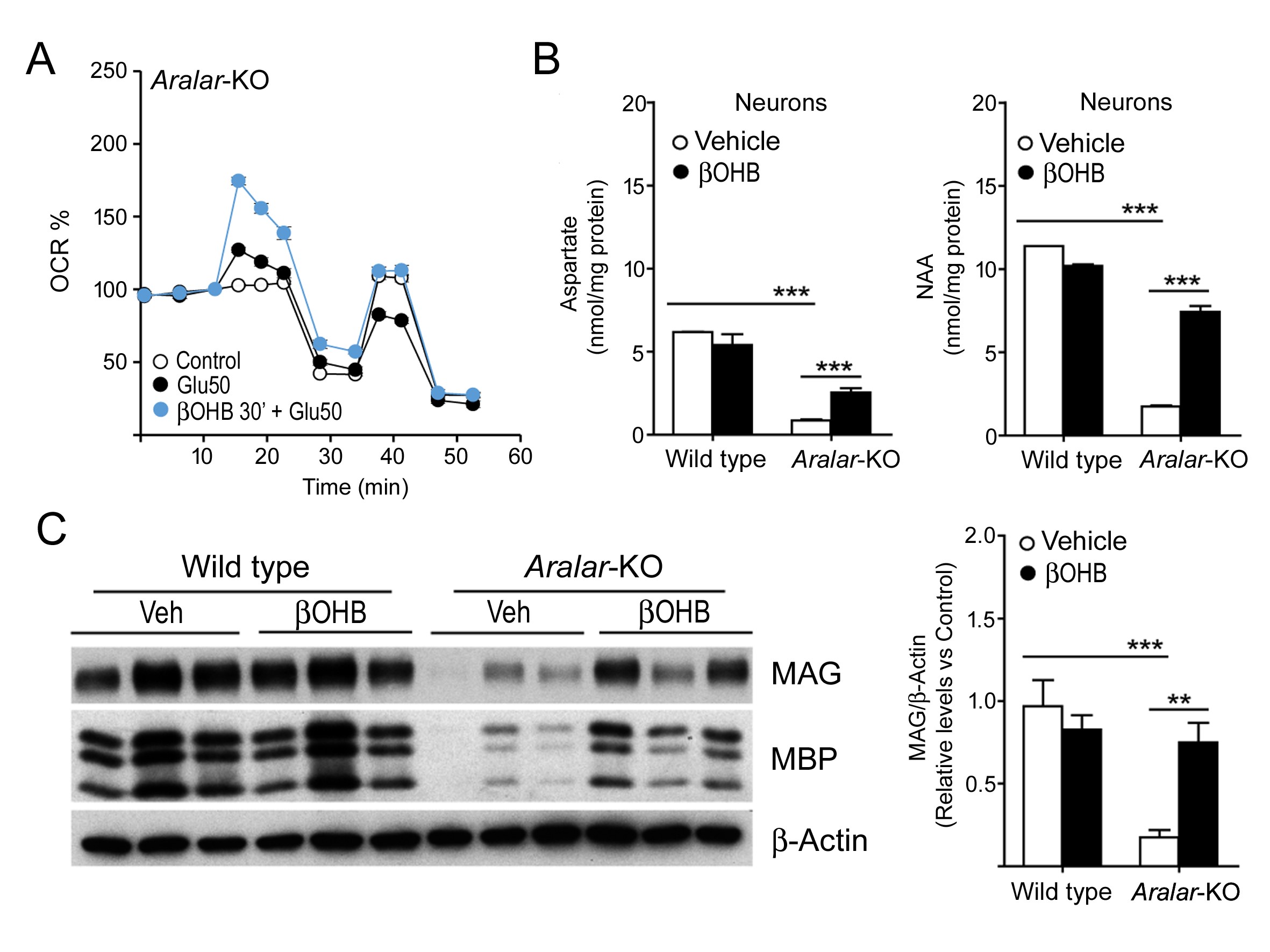
Figura. Efectos protectores de βOHB en neuronas y cerebro de ratones Aralar-KO: una alternativa a la dieta cetogénica. βOHB incrementa la respiración estimulada por glutamato (A) y el contenido de Aspartato y NAA (B) en cultivos de neuronas Aralar-KO, y los niveles de proteínas de la mielina en regiones de la corteza en ratones Aralar-KO (C) tras su administración in vivo.

| Apellidos | Nombre | Laboratorio | Ext.* | Categoría profesional | |
|---|---|---|---|---|---|
| Bogónez Peláez | Elena | 321 | 4622 | ebogonez(at)cbm.csic.es | Profesor Titular Universidad, GA |
| Carrascosa Baeza | José Mª | 321 | 4651 | jmcarrascosa(at)cbm.csic.es | Catedrático Universidad, GA |
| Contreras Balsa | Laura | 321 | 4651 | lcontreras(at)cbm.csic.es | Titulado Sup.de Actividades Técn. y Profes. GP1 |
| Del Arco | Araceli | 321 | 4651 | adelarco(at)cbm.csic.es | Prof Titular Universidad (convenio UAM-UCLM) |
| González Moreno | Luis | 321 | 4651 | lgmoreno(at)cbm.csic.es | Contrato Predoctoral |
| Juaristi Santos | Inés | 321 | 4651 | ijuaristi(at)cbm.csic.es | Investigador |
| Pardo Merino | Beatriz | 321 | 4635 | bpardo(at)cbm.csic.es | Profesor Titular Universidad, GA |
| Richter | Emma | 321 | 4651 | Becario Erasmus | |
| Romeral Buzón | Alejandro | 4651 | Estudiante TFG | ||
| Santamaría Cano | Andrea | 321 | 4651 | asantamaria(at)cbm.csic.es | Contrato Predoctoral |
| Satrústegui Gil-Delgado | Jorgina | 321 | 4621 | jsatrustegui(at)cbm.csic.es | Emérito |
| Sesé Cobos | Bárbara | 321 | 4651 | bsese(at)cbm.csic.es | E.Ayudantes De Invest. De Los Oo.Publicos De Investigacion |
| Von-Kobbe Alonso | Cayetano-Alfonso | 114.4 | 4809 | cvonkobbe(at)cbm.csic.es |
E.Científicos Titulares de Organismos Públicos de Investigación |
* For external calls please dial 34 91196 followed by the extension number
Publicaciones relevantes:
- Pérez-Liébana I, Juaristi I, González-Sánchez P, González-Moreno L, Rial E, Podunavac M, Zakarian A, Molgó J, Vallejo-Illarramendi A, Mosqueira-Martín L, Lopez de Munain A, Pardo B, Satrústegui J, Del Arco A. A Ca2+-Dependent Mechanism Boosting Glycolysis and OXPHOS by Activating Aralar-Malate-Aspartate Shuttle, upon Neuronal Stimulation. J Neurosci. 2022 May 11;42(19):3879-3895. doi: 10.1523/JNEUROSCI.1463-21.2022. Epub 2022 Apr 6. PMID: 35387872; PMCID: PMC9097769.
- Sánchez-González C, Herrero Martín JC, Salegi Ansa B, Núñez de Arenas C, Stančič B, Pereira MP, Contreras L, Cuezva JM, Formentini L. Chronic inhibition of the mitochondrial ATP synthase in skeletal muscle triggers sarcoplasmic reticulum distress and tubular aggregates. Cell Death Dis. 2022 Jun 22;13(6):561. doi: 10.1038/s41419-022-05016-z. PMID: 35732639; PMCID: PMC9217934.
- Pardo B, Herrada-Soler E, Satrústegui J, Contreras L, Del Arco A. AGC1 Deficiency: Pathology and Molecular and Cellular Mechanisms of the Disease. Int J Mol Sci. 2022 Jan 4;23(1):528. doi: 10.3390/ijms23010528. PMID: 35008954; PMCID: PMC8745132.
- Nuevo-Tapioles C, Santacatterina F, Sánchez-Garrido B, de Arenas CN, Robledo-Bérgamo A, Martínez-Valero P, Cantarero L, Pardo B, Hoenicka J, Murphy MP, Satrústegui J, Palau F, Cuezva JM. Effective therapeutic strategies in a preclinical mouse model of Charcot-Marie-Tooth disease. Hum Mol Genet. 2021 Nov 30;30(24):2441-2455. doi: 10.1093/hmg/ddab207. PMID: 34274972; PMCID: PMC8643506.
- Pérez-Liébana I, Juaristi I, González-Sánchez P, González-Moreno L, Rial E, Podunavac M, Zakarian A, Molgó J, Vallejo-Illarramendi A, Mosqueira-Martín L, Lopez de Munain A, Pardo B, Satrústegui J, Del Arco A. A Ca2+-Dependent Mechanism Boosting Glycolysis and OXPHOS by Activating Aralar-Malate-Aspartate Shuttle, upon Neuronal Stimulation. J Neurosci. 2022 May 11;42(19):3879-3895. doi: 10.1523/JNEUROSCI.1463-21.2022. Epub 2022 Apr 6. PMID: 35387872; PMCID: PMC9097769.
- Sánchez-González C, Herrero Martín JC, Salegi Ansa B, Núñez de Arenas C, Stančič B, Pereira MP, Contreras L, Cuezva JM, Formentini L. Chronic inhibition of the mitochondrial ATP synthase in skeletal muscle triggers sarcoplasmic reticulum distress and tubular aggregates. Cell Death Dis. 2022 Jun 22;13(6):561. doi: 10.1038/s41419-022-05016-z. PMID: 35732639; PMCID: PMC9217934.
- J, Contreras L, Del Arco A. AGC1 Deficiency: Pathology and Molecular and Cellular Mechanisms of the Disease. Int J Mol Sci. 2022 Jan 4;23(1):528. doi: 10.3390/ijms23010528. PMID: 35008954; PMCID: PMC8745132.
- Nuevo-Tapioles C, Santacatterina F, Sánchez-Garrido B, de Arenas CN, Robledo-Bérgamo A, Martínez-Valero P, Cantarero L, Pardo B, Hoenicka J, Murphy MP, Satrústegui J, Palau F, Cuezva JM. Effective therapeutic strategies in a preclinical mouse model of Charcot-Marie-Tooth disease. Hum Mol Genet. 2021 Nov 30;30(24):2441-2455. doi: 10.1093/hmg/ddab207. PMID: 34274972; PMCID: PMC8643506.
- Lunetti P, Marsano RM, Curcio R, Dolce V, Fiermonte G, Cappello AR, Marra F, Moschetti R, Li Y, Aiello D, Del Arco Martínez A, Lauria G, De Leonardis F, Ferramosca A, Zara V, Capobianco L. (2021) The mitochondrial aspartate/glutamate carrier (AGC or Aralar1) isoforms in D. melanogaster: biochemical characterization, gene structure, and evolutionary analysis. Biochim Biophys Acta Gen Subj. 1865:129854.
- Esparza-Moltó PB, Romero-Carramiñana I, Núñez de Arenas C, Pereira MP, Blanco N, Pardo B, Bates GR, Sánchez-Castillo C, Artuch R, Murphy MP, Esteban JA, Cuezva JM. (2021) Generation of mitochondrial reactive oxygen species is controlled by ATPase inhibitory factor 1 and regulates cognition. PLoS Biol. 19:e3001252.
- García-Catalán S, González-Moreno L, Del Arco A. (2021) Ca2+-regulated mitochondrial carriers of ATP-Mg2+/Pi: Evolutionary insights in protozoans. Biochim Biophys Acta Mol Cell Res. 1868:119038.
- Rubio C, Lizárraga E, Álvarez-Cilleros D, Pérez-Pardo P, Sanmartín-Salinas P, Toledo-Lobo MV, Alvarez C, Escrivá F, Fernández-Lobato M, Guijarro LG, Valverde AM, Carrascosa JM. (2020) Aging in male Wistar rats associates with changes in intestinal microbiota, gut structure and cholecystokinin-mediated gut-brain axis function. J Gerontol A Biol Sci Med Sci. 2020 Dec 14:glaa313. doi: 10.1093/gerona/glaa313.
- Civera-Tregón A, Domínguez L, Martínez-Valero P, Serrano C, Vallmitjana A, Benítez R, Hoenicka J, Satrústegui J, Palau F. (2021) Mitochondria and calcium defects correlate with axonal dysfunction in GDAP1-related Charcot-Marie-Tooth mouse model. Neurobiol Dis. 152:105300. doi: 10.1016/j.nbd.2021.
- Pérez-Liébana I, Casarejos MJ, Alcaide A, Herrada-Soler E, Llorente-Folch I, Contreras L, Satrústegui J, Pardo B. (2020) βOHB Protective Pathways in Aralar-KO Neurons and Brain: An Alternative to Ketogenic Diet. J Neurosci. 40:9293-9305. doi: 10.1523/JNEUROSCI.0711-20.2020
- Rubio C, Puerto M, García-Rodríquez JJ, Lu VB, García-Martínez I, Alén R, Sanmartín-Salinas P, Toledo-Lobo MV, Saiz J, Ruperez J, Barbas C, Menchén L, Gribble FM, Reimann F, Guijarro LG, Carrascosa JM, Valverde ÁM. (2020) Impact of global PTP1B deficiency on the gut barrier permeability during NASH in mice. Mol Metab. 35:100954. doi: 10.1016/j.molmet.2020.01.018
- Juaristi I, Llorente-Folch I, Satrústegui J, Del Arco A. (2019) Extracellular ATP and glutamate drive pyruvate production and energy demand to regulate mitochondrial respiration in astrocytes. Glia 67:759-774. doi: 10.1002/glia.23574.
- Puertas-Frías G, Del Arco A, Pardo B, Satrústegui J, Contreras L (2019) Mitochondrial movement in Aralar/Slc25a12/AGC1 deficient cortical neurons. Int. 131:104541. doi: 10.1016/j.neuint.2019.104541.
- von Kobbe C (2019) Targeting senescent cells: approaches, opportunities, challenges. Aging (Albany NY) 11: 12844–12861. doi: 10.18632/aging.102557.
- Remacha L, Comino-Méndez I, Richter S, Contreras L, Currás-Freixes M, Pita G, Letón R, Galarreta A, Torres-Pérez R, Honrado E, Jiménez S, Maestre L, Moran S, Esteller M, Satrústegui J, Eisenhofer G, Robledo M, Cascón A. (2017) Targeted Exome Sequencing of Krebs Cycle Genes Reveals Candidate Cancer-Predisposing Mutations in Pheochromocytomas and Paragangliomas. Clin Cancer Res. 23:6315-6324. doi: 10.1158/1078-0432.CCR-16-2250.
- González-Sánchez P, Del Arco A, Esteban JA, Satrústegui J. (2017) Store-Operated Calcium Entry Is Required for mGluR-Dependent Long Term Depression in Cortical Neurons. Front Cell Neurosci. 11:363. doi: 10.3389/fncel.2017.00363.
- Writzl K, Maver A, Kovačič L, Martinez-Valero P, Contreras L, Satrustegui J, Castori M, Faivre L, Lapunzina P, van Kuilenburg ABP, Radović S, Thauvin-Robinet C, Peterlin B, Del Arco A, Hennekam RC. (2017) De Novo Mutations in SLC25A24 Cause a Disorder Characterized by Early Aging, Bone Dysplasia, Characteristic Face, and Early Demise. Am J Hum Genet. 101:844-855. doi: 10.1016/j.ajhg.2017.09.017.
- Juaristi I, García-Martín ML, Rodrigues TB, Satrústegui J, Llorente-Folch I, Pardo B. (2017) ARALAR/AGC1 deficiency, a neurodevelopmental disorder with severe impairment of neuronal mitochondrial respiration, does not produce a primary increase in brain lactate. J Neurochem. 142:132-139. doi: 10.1111/jnc.14047.
- González-Sánchez P, Pla-Martín D, Martínez-Valero P, Rueda CB, Calpena E, Del Arco A, Palau F, Satrústegui J. (2017) CMT-linked loss-of-function mutations in GDAP1 impair store-operated Ca2+ entry-stimulated respiration. Sci Rep. 7:42993. doi: 10.1038/srep42993.
- Rueda, C.B., Traba, J., Amigo, I., Llorente-Folch, I., González-Sánchez, P., Pardo, B., Esteban, J.A., del Arco, A. and Satrústegui, J. (2015) Mitochondrial ATP-Mg/Pi carrier SCaMC-3/Slc25a23 counteracts PARP-1- dependent fall in mitochondrial ATP caused by excitotoxic insults in neurons. J. Neurosci. 35, 3566-3581.
- Llorente-Folch, I., Rueda, C.B., Pardo, B., Szabadkai, G., Duchen, M.R. and Satrustegui, J. (2015) The regulation of neuronal mitochondrial metabolism by calcium. J. Physiol. 593, 3447-3462.
- Llorente-Folch, I., Rueda, C.B., Perez-Liebana, I., Satrústegui, J. and Pardo, B. (2016) L-Lactate-Mediated Neuroprotection against Glutamate-Induced Excitotoxicity Requires Aralar/AGC1. J. Neurosci. 36, 4443-4456.
- Amigo, I., Traba, J., González-Barroso, MM., Rueda, CB., Fernández, M., Rial, E., Sánchez, A, Satrústegui, J. and del Arco, A. (2013) Glucagon regulation of oxidative phosphorylation requires an increase in matrix adenine nucleotide content through Ca2+ activation of the mitochondrial ATP-Mg/Pi carrier SCaMC-3. J. Biol. Chem. 288, 7791-7802.
- Llorente-Folch, I., Rueda, CB., Amigo, I., del Arco, A., Saheki, T., Pardo, B. and Satrústegui, J. (2013) Calcium-regulation of mitochondrial respiration maintains ATP homeostasis and requires ARALAR/AGC1-malate aspartate shuttle in intact cortical neurons. J. Neurosci. 33, 13957-13971.
- Lindsay, KJ., Du, J., Sloat, SR., Contreras, L., Linton, JD., Turner, SJ., Sadilek, M., Satrústegui, J. and Hurley, JB. (2014) Pyruvate kinase and aspartate-glutamate carrier distributions reveal key metabolic links between neurons and glia in retina. Proc. Natl. Acad. Sci. U. S. A. 111, 15579-15584.
- Llorente-Folch, I., Sahún, I., Contreras, L., Casarejos, MJ., Grau JM, Saheki, T., Mena, MA., Satrústegui, J., Dierssen M. and Pardo, B. (2013) AGC1-malate aspartate shuttle activity is critical for dopamine handling in the nigrostriatal pathway. J. Neurochem. 124, 347-362.
Capítulos de libros:
- del Arco, A. and Satrústegui, J. (March 2013) Mitochondrial Carriers. In: eLS. John Wiley & Sons, Ltd: Chichester.
- José María Carrascosa Baeza, “Ciencia, ética y sostenibilidad: el papel de la Academia para alcanzar los Objetivos de Desarrollo Sostenible” Agenda 2030: Claves para la transformación sostenible / Margarita Alfaro (ed. lit.), Silvia Arias Careaga (ed. lit.), Ana Gamba Romero (ed. lit.), ISBN 9788490977071.
Tesis doctorales:
- Irene Pérez Liébana (2020) “The mitochondrial aspartate-glutamate carrier Aralar/AGC1 controls neuronal respiration and (beta)-Hydroxybutyrate rescues brain defects caused by AGC1 deficiency”. UAM. Jorgina Satrústegui y Beatriz Pardo.
- Inés Juaristi Santos (2019) “Regulation of mitochondrial respiration in astrocytes: role of Ca2+, ATP demand and pyruvate production”. UAM. Jorgina Satrústegui y Araceli del Arco.
- Carmen Rubio Caballero (2018) “Changes in gut microbiota associated with inflammation during ageing and non-alcoholic steatohepatitis”. UAM. José María Carrascosa Baeza y Ángela Martínez Valverde.
- Paula Martínez Valero (2017) “Enfermedad de Charcot-Marie-Tooth asociada a Gadp1: alteraciones en nocicepción y señalización por calcio”. UAM. Jorgina Satrústegui, Laura Contreras y Carolina Roza.
- Paloma González Sánchez (2017) “Store-operated calcium entry in neural cells. A role in Charcot-Marie-Tooth disease”. UAM. Jorgina Satrústegui y Araceli del Arco.
- Carlos B. Rueda Díez (2014). “Ca2+ modulation of mitochondrial function under physiological and pathological stimulation: Role of the ATP-Mg/Pi carrier, SCaMC-3”. UAM. Jorgina Satrústegui, Beatriz Pardo y Araceli del Arco.
- Irene Llorente Folch (2013). “New roles of aralar, the brain mitochondrial aspartate/glutamate carrier in dopamine handling glutamate excitotoxicity and regulation of mitochondrial respiration”. UAM. Jorgina Satrústegui y Beatriz Pardo.
- Ignacio Amigo de la Huerga (2013). “Characterization of SCaMC-3, the mitochondrial ATP-Mg/Pi carrier present in liver and brain”. UAM. Jorgina Satrústegui y Araceli del Arco


















