Programa Científico
Homeostasis de tejidos y órganos
UNIDADES EN ESTE PROGRAMA
Arquitectura celular y organogénesis Comunicación intercelular e inflamación
GRUPO DE INVESTIGACIÓN
Mecanismos de tumorigénesis en Drosophila

Estamos interesados en el estudio de la senescencia celular, un fenómeno involucrado en procesos inflamatorios y tumorogénesis descrito en vertebrados.
Recientemente hemos encontrado que las células senescentes de Drosophila tienen las mismas características que las de vertebrados. Empleamos la sofisticada genética desarrollada en Drosophila para describir las modificaciones genéticas que sufren las células senescentes y las interacciones con sus vecinas.

Investigación
El principal objetivo de nuestra investigación es el estudio de la formación de tumores, un fenómeno de máxima relevancia tanto científica como social. Debido a su sofisticada tecnología genética, así como otras ventajas a nivel experimental, usamos los discos imaginales de la mosca de la fruta, Drosophila melanogaster, como sistema modelo para investigar los factores moleculares y genéticos que subyacen a la aparición de tumores. Nuestro trabajo se centra en tres líneas de investigación.
1)El potencial tumorigénico de células que no pueden ejecutar el programa apoptótico.
Esta línea se basa en trabajos previos del laboratorio sobre el comportamiento de células en las cuales la maquinaria apoptótica ha sido suprimida; estas células deficientes para la apoptosis (Apoptosis-Deficient, AD) sobreviven al estrés o a los tratamientos con drogas que conducen a la muerte a las células normales. Hemos desarrollado un modelo de tumorogénesis basado en las propiedades de estas células: tras inducir un estrés transitorio una fracción de la población de células AD activa ectópicamente las funciones pro-proliferativas y pro-migratorias de la vía de la Jun N-Terminal Kinase (JNK), lo cual resulta en la generación de tumores invasivos. La generación de estos tumores está mediada por la actividad de la JNK. Es digna de mención la observación de que la actividad inapropiada de la vía de la JNK está asociada con tumores en humanos.

Basándonos en que JNK parece ser un factor determinante en el efecto oncogénico, estamos estudiando los mecanismos por los cuales las células AD que expresan JNK inducen la formación de tumores (figura 1). Hemos visto que presentan características de células senescentes, incluyendo el fenotipo secretor asociado a la senescencia (SASP). En particular, éstas células secretan moléculas señalizadoras como Dpp, Wg y Upd encargadas de activar las rutas de Dpp, Wg y JAK/STAT respectivamente. Experimentos en los cuales se han suprimido estas vías indican que su función es responsable de los sobrecrecimientos tumorales, aunque se desconocen los efectos específicos de cada una de las vías así como las interacciones entre ellas que están siendo objeto de estudio en el laboratorio. Tenemos evidencias preliminares de que no todas las células senescentes adquieren el fenotipo SASP; aquellas con SASP aparecen en la región central del disco, mientras que las localizadas en el notum o en regiones periféricas no presentan SASP. La observación de que existen células senescentes SASP y no-SASP es nueva en el campo. Correlaciona con la observación de que los crecimientos tumorales ocurren en la región central del disco, lugar dónde se concentran las células senescentes secretoras. El mecanismo por el cual se produce este efecto dependiente de posición está siendo investigado en el laboratorio.
Un aspecto relacionado es cómo la actividad de la JNK es capaz de ejercer todas estas funciones. Hace unos años llevamos a cabo una búsqueda por RNAseq de genes específicamente activados por la vía de la JNK y necesarios para generar sobrecrecimientos. Algunos de los genes identificados están siendo investigados por su implicación con las señales que emanan de las células senescentes y los sobrecimientos asociados.
2) Modificaciones de la cromatina en células senescentes y reversibilidad de la senescencia
La senescencia puede ser inducida por uno o varios estímulos que producen estrés celular (como, por ejemplo, daño al ADN, estrés oxidativo o activación de oncogenes). Todos ellos producen alteraciones, generalmente estables, en la función y estructura de la célula. Uno de los marcadores de la transformación es un cambio en la estructura de la cromatina, denominado “senescence-associated heterochromatin foci (SAHFs)” es decir, focos de heterocromatina asociada a senescencia. En células de mamíferos, tras la inducción de senescencia aparecen los SAHFs como focos enriquecidos en marcadores de heterocromatina (como H3K9me3) y con niveles disminuidos de marcadores eucromáticos. En el laboratorio estamos empezando a describir el proceso por el cual las células entran en senescencia, algo que no ha sido descrito hasta la fecha en Drosophila. Hemos visto que, análogamente a lo que ocurre en células de vertebrados en cultivo, las células del disco de ala de Drosophila aumentan el tamaño de sus nucléolos al volverse senescentes. Los niveles de H3K9me3 también aumentan en esas células. Dado que la vía de la JNK es un inductor de senescencia, estamos estudiando cómo actúa sobre las modificaciones en cromatina. Algunas de las dianas de JNK identificadas en la búsqueda de ARNs mencionada anteriormente están asociadas con modificaciones cromatínicas (como Caf1-105, involucrado en la formación y el mantenimiento de la heterocromatina). Estamos analizando las posibles implicaciones de algunos de esos genes en la remodelación de la cromatina asociada con la senescencia.
Además, hemos encontrado un función inesperada de los transposones F-element (pertenecientes a la familia LINE) en el mantenimiento del estado de senescencia; al expresarlos a altos niveles las células senescentes parecen revertir a su condición silvestre (figura 2). Se trata de una observación de gran interés, en primer lugar porque sugiere que estos transposones-que ya se ha visto que tienen funciones en la remodelación de la cromatina durante los primeros estadios embrionarios tanto en vertebrados como en Drosophila -funcionan también en senescencia, y en segundo lugar porque es el primer indicio descrito de que la senescencia puede ser revertida. Los detalles moleculares de este fenómeno están actualmente siendo investigados.
3) Interacciones entre poblaciones celulares con distintas tasas de proliferación
En nuestros análisis de tumores producidos por mutaciones oncogénicas en polyhomeotic ((ph)) y erupted (el Drosophila homólogo en Drosophila del gen tumor susceptibility gene 101), hemos realizado observaciones que pudieran tener implicaciones a la hora de entender cómo las células tumorales colonizan tejidos adyacentes: en experimentos en los cuales un compartimento del disco de ala es silvestre y el otro sobrecrece debido a que es mutante para el oncogen ph o erupted el compartimento silvestre crece menos que su tamaño en condiciones normales
Hemos observado que este fenómeno ocurre también entre regiones del mismo compartimento. Lo que parece estar ocurriendo es que la población de células que sobrecrece inhibe el crecimiento de los tejidos vecinos. Nos referimos a esas poblaciones como “ganadora” (la que sobrecrece) y “perdedora” (la que pierde tamaño). Hemos descartado la hipótesis de que la población perdedora sufra un incremento de muerte por apoptosis, pero hemos visto que esta población tiene un índice mitótico menor de lo normal, lo cual explica su crecimiento reducido. Dado que el crecimiento general del disco depende de factores de crecimiento señalizadores como Dpp o Wg, estamos estudiando la posibilidad de que las células de la población ganadora secuestren las moléculas señalizadoras Dpp o Wg impidiendo su accesiblidad a la población perdedora.
También estamos tratando de identificar los factores genéticos asociados con la reducción del índice mitótico, examinando los niveles de expresión de genes reguladores de la división celular, tanto los que la promueven (cdk, y otras quinasas dependientes de ciclinas) como las que la inhiben (dacapo, frs). Existe también la posibilidad de que la via de Hippo, un inhibidor de la proliferación celular, esté alterada, lo cual podría testarse usando numerosos marcadores disponibles para monitorizar su actividad (expanded, warts, Yki).
Miembros del grupo

Manuel Calleja Requena
Lab.: 422.1 Ext.: 4697/4683
mcalleja(at)cbm.csic.es

Natalia Azpiazu Torres
Lab.: 422 Ext.: 4697
nazpiazu(at)cbm.csic.es

Ginés Morata Pérez
Lab.: 422 Ext.: 4696
gmorata(at)cbm.csic.es

Angélica Cantarero Mateo
Lab.: 422 Ext.: 4607/4683
acantarero(at)cbm.csic.es

David Foronda Alvaro
Lab.: 422 Ext.: 4699
dforonda(at)cbm.csic.es

Noelia Pinal Seoane
Lab.: 422 Ext.: 4683
npinal(at)cbm.csic.es

Catalina Grabowski Pinto
Lab.: 422 Ext.: 4697
cgrabowski(at)cbm.csic.es
Silvia Vega Esquinas
Lab.: 422 Ext.: 4697
silvia.vega(at)cbm.csic.es
Carmen Lucía Jarabo Caruda
Lab.: 422 Ext.: 4683
Eduardo García Cárdaba
Lab.: 422 Ext.: 4697
Publicaciones representativas
Short-term activation of the Jun N-terminal kinase pathway in apoptosis-deficient cells of Drosophila induces tumorigenesis
Noelia Pinal et al.
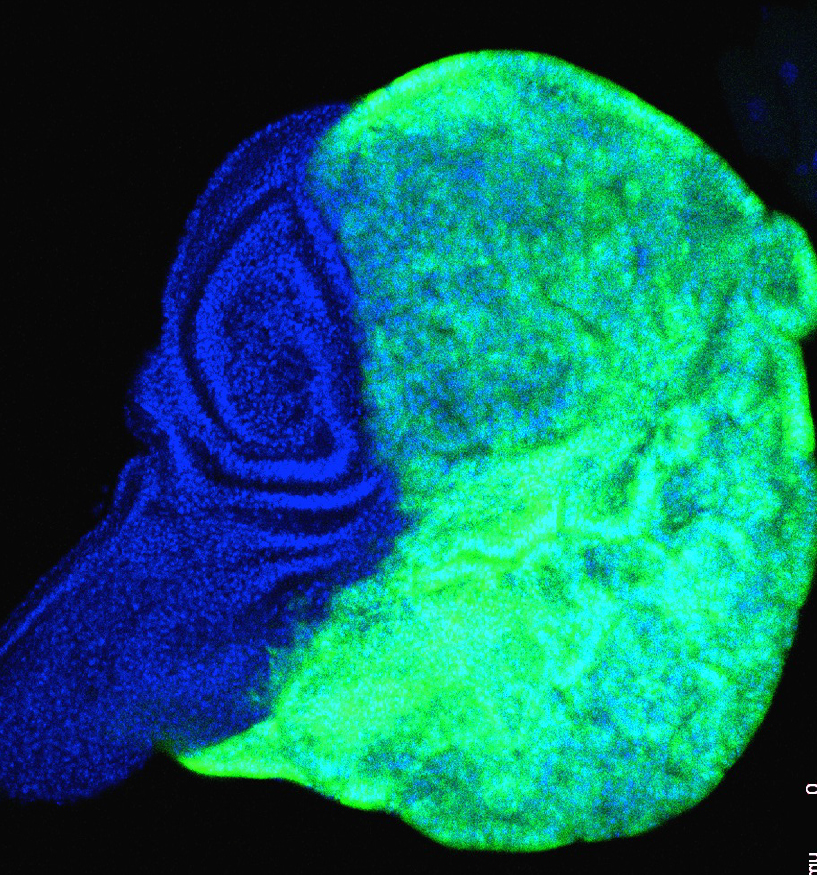
Tumorigenesis and cell competition in Drosophila in the absence of polyhomeotic function
Izarne Medina et al.

Chromatin remodelling and retrotransposons activities during regeneration in Drosophila
Natalia Azpiazu et al.

Lack of apoptosis leads to cellular senescence and tumorigenesis in Drosophila epithelial cells
Juan Manuel Garcia-Arias et al.
Últimas publicaciones
Programas Científicos